
Géomorphologie et hydrologie karstiques
 |
A travers sa thèse, soutenue en 2009, portant sur la géologie structurale de la chaîne de Lengguru, V. Bailly (1) montre que les massifs karstiques sont issus de la compression et de la déformation d’un prisme d’accrétion suite à la subduction de la plaque pacifique sous la plaque australienne. Cet événement géologique majeur est daté de près de 10 millions d’années. L’étude a également révélé que le massif s’effondre depuis environ 3 millions d’années. Ce processus est matérialisé par l’apparition de nombreuses fractures perpendiculaires aux plis de compression ce qui pourrait avoir un impact majeur sur le réseau hydrographique karstique et notamment sur la dynamique hydrologique des lacs situés dans les poljés. Une attention particulière sera portée sur le fonctionnement hydrologique de ces lacs et notamment sur la fluctuation de leur niveau.(2)
Les ressources hydrologiques, en quantité (débits) et qualité (géochimie), sont la clef de voute de l’équilibre des écosystèmes karstiques et de leur biodiversité associée. Il est donc primordial de caractériser le contexte géomorphologique et hydrologique des karsts de Lengguru puis de présager de l’évolution future de ces ressources par l’utilisation de modèles prédictifs basés sur des situations évolutives contrastées. Dans ce contexte, cinq zones cibles ont été choisies dans différentes parties du massif grâce à la superposition de données satellitaires, de cartographie et d’imagerie aériennes. Elles ont été définies à proximité des principaux lacs de la région et se situent dans des parties très érodées du karst, matérialisées par des dolines, des gouffres et de nombreuses disparitions-résurgences de rivières.
Un bilan hydrologique lacustre incluant une estimation des apports et des pertes du réseau karstique, sera effectué sur les lacs Aiwasa, Lakamora, Kamaka, Sewiki et Mbuta.
L’exploration spéléologique complètera le bilan hydrologique lacustre et permettra notamment la cartographie, la topographie et l’étude des mécanismes de dépôts, d’érosion et de concrétion qui caractérisent ces réseaux souterrains.■
|



(1) Thèse de Vivien Bailly: « La chaîne de Lengguru: Evolution et structure d’un prisme jeune dans le contexte tectonique rapide de Papouasie occidentale. »
Soutenue le 13 novembre 2009 pour l’obtention du titre de Docteur de l’Université Paris VI -Spécialité Sciences de la Terre, Ecole Doctorale GRN ED 398 préparée au laboratoire de Géologie de l’Ecole Normale Supérieure de Paris et dans les locaux de Total E&P Indonésie Jakarta.
(2) Article 2008 Bailly, V., Pubellier M., Ringenbach J.C., Structure of the Lengguru Fold Belt, New Guinea island : consequence of rapid kinematic changes 2008, 33 International Geological Congress, Oslo.
Paléontologie : les écosystèmes disparus
Le travail de thèse de V. Bailly signale dans le massif karstique et à ses abords différentes formations et ensembles sédimentaires potentiellement fossilifères.
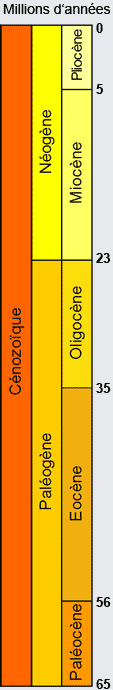
Il en décrit les principaux caractères. Ainsi nous savons que dans notre zone d’intérêt, quatre formations et ensembles sédimentaires sont présents et datent tous du Cénozoïque. Il s’agit : de l’ensemble Paléogène (23 à 65 millions d’années) et Miocène (5 à 23 millions d’années ) calcaire qui constitue le massif karstique lui-même où divers faciès ont été identifiés et caractérisés entre autres par des associations de foraminifères, de gastéropodes et de bivalves ; d’une formation Mio-Pliocène typiquement syn-tectonique comme l’illustre la présence de slumps (pli créé par gravité dans un sédiment non-consolidé) qui contient non seulement des microfossiles mais aussi des fossiles de vertébrés ; d’une formation Plio-Pléistocène considérée post-tectonique (d’accrétion), constituée de boues calcaires et de bancs à dominantes détritiques, incluant des coquilles de mollusques et localement des débris végétaux ; et de dépôts du Quaternaire semblables aux dépôts actuels et incluant des bio-constructions en zone côtière.

La collecte des informations paléontologiques, essentiellement des fossiles d’organismes mais aussi des traces, va viser deux objectifs indépendants : d’une part renseigner précisément les environnements passés au cours de la mise en place et de l’évolution du massif karstique, et d’autre part, documenter l’évolution des espèces qui l’ont peuplé tout au long de ce processus. Pour cette raison, nous allons donc nous concentrer sur le contenu fossilifère des formations néogènes (23 millions d’années à nos jours).

Les fossiles sont de bons outils de reconstitution des paléo-environnements soit en tant que marqueurs biologiques de la niche qu’ils occupaient, soit en tant qu’«enregistreurs», dans leurs formations squelettiques, des données géochimiques de l’environnement avec par exemple l’isotope 18 de l’atome d’oxygène ou δ18O. Les microfossiles en particulier donnent une information directe sur le milieu de dépôt, marin ou dulçaquicole, pour les zones qui ont pu subir alternativement l’une ou l’autre des influences. Ils permettent éventuellement de dater leur niveau par comparaison avec des échelles bio-stratigraphiques de référence. Des échantillonnages systématiques par carottage seront effectués dans les cinq zones d’intérêt, avec une attention particulière pour les dépôts de surface des lacs et les remplissages sédimentaires de poches souterraines. L’objectif est de pouvoir identifier d’éventuelles séquences d’assèchement dans les dépôts récents, d’en caractériser la dynamique bio-sédimentaire, et de tenter de reconnaitre un éventuel contrôle soit par les pulsations climatiques du Plio-Quaternaire soit par les phases microtectoniques locales récentes en distension.

Mesure de pendage
En plus de leur efficacité dans la reconstitution des paléo-environnements, les fossiles nous intéressent aussi pour les informations uniques qu’ils apportent sur l’évolution des groupes. Effectivement, seule leur étude permet –particulièrement chez les vertébrés- de connaitre des caractères ou des associations de caractères morphologiques qui n’existent plus chez les espèces actuelles. Par ailleurs, les fossiles permettent de reconstituer des distributions passées plus larges ou différentes de ce que laissent présager les seuls représentants actuels d’un groupe. A ce titre, les fossiles des groupes étudiés dans l’actuel (les poissons, les gastéropodes, les crustacés et les insectes) seront activement recherchés dans les trois formations néogènes. Dès à présent nous sommes assurés de trouver au moins des fossiles de poissons et de gastéropodes qui ont peuplé la région ces derniers millions d’années. Leur identification ouvrira une fenêtre sur des peuplements disparus de l’île et plus précisément sur leurs éventuels liens avec les occupants actuels, permettant alors une calibration des arbres moléculaires indépendante du contexte géologique.
Biodiversité exceptionnelle
Du fait de leur configuration structurale et des importantes réserves en eaux qu’ils contiennent, les massifs karstiques abritent une biodiversité importante aussi bien dans les écosystèmes de surface que souterrains. Effectivement, l’intense activité géomorphologique qui caractérise ces massifs conduit souvent à la fragmentation des habitats et à l’apparition d’écosystèmes très originaux. Ces mécanismes sont généralement promoteurs de spéciation et font des systèmes karstiques de véritables « usines à espèces », en particulier lorsqu’ils montrent une évolution géomorphologique intense.
La fragmentation extrême des habitats dans la région de Kaimana a permis une radiation remarquable du groupe des
poissons arc-en-ciel. A travers les différents spécialistes impliqués dans cette campagne scientifique, d’autres groupes –
mollusques, insectes, crustacés au fort potentiel de découvertes seront aussi étudiés.
Le contexte de surrection rapide de la chaine de Lengguru, associé à une intense activité d’érosion, a conduit ces quelques derniers millions d’années à la formation d’un massif très fragmenté et constitué de multiples dômes carbonatés séparés par des dépressions (poljés) qui peuvent selon leur altitude être ennoyées par la mer ou former de vastes lacs ou marécages. Ce massif doit donc contenir une diversité exceptionnelle, en particulier d’espèces très spécialisées et présentant des capacités limitées de dispersion.
Cette expédition se focalisera sur plusieurs groupes zoologiques incluant la microfaune dans son ensemble, ainsi que les poissons, les gastéropodes hydrobies, les crustacés et les insectes. Ces modèles biologiques seront échantillonnés dans les cinq zones cibles définies par les hydrogéologues, et ce, aussi bien dans les écosystèmes de surface que dans les parties souterraines. Il est prévu de s’intéresser à d’autres modèles biologiques (plantes, oiseaux, petits mammifères, arachnides, reptiles) dans le cadre d’expéditions futures, l’échantillonnage requérant des dispositifs dédiés dont la mise en œuvre peut être longue.

Figure 5 – Pour accéder aux sites d’étude, les explorateurs de l’expédition devront parcourir à pied ou en bateau les nombreux cours
d’eau du massif (A) afin d’y installer les camps de base (B) véritables centres de recherches et de vie (C). Par ailleurs, le projet
Lengguru-Kaimana 2010 inclut le tournage d’un film documentaire de 52′, à destination du grand public, une coproduction IRD-GEDEON.
Un inventaire exhaustif des poissons, incluant d’éventuelles espèces souterraines, sera effectué dans les zones étudiées avec une attention particulière pour les groupes très diversifiés des poissons arc-en-ciel (Melanotaeniidae) et des espèces diadromes amphidromes (Gobiidae, Eleotridae). Dans l’état actuel des connaissances, six espèces de poissons arc-en-ciel, dont cinq endémiques (genres Melanotaenia et Pelangia) sont décrites des lacs et rivières bordant le massif karstique de Lengguru. Il n’y a par contre aucune donnée dans les parties isolées du vaste et complexe réseau hydrologique au cœur du massif. L’objectif sera d’une part de récolter toutes ces espèces nominales pour compléter les analyses phylogénétiques d’une thèse en cours (thèse de Doctorat de M. Kadarusman, UR CAVIAR, UPS) et d’autre part d’explorer des lacs et rivières isolés pour y découvrir d’éventuelles espèces nouvelles. Cet inventaire, au delà de son intérêt propre, nourrira l’étude des processus évolutifs de diversification qui ont façonné l’ichtyofaune, de les replacer dans le contexte régional et de prédire l’évolution future des populations les plus exposées aux activités humaines et aux modifications d’habitats.

Les systèmes insulaires de l’Indopacifique abritent des espèces diadromes (cycle de vie partagé entre les eaux marine et douce) qui sont les seules espèces capables de coloniser naturellement les cours d’eau. Leur stratégie de dispersion représente un moteur essentiel de la structuration et de la persistance des communautés. Les espèces diadromes amphidromes (Gobiidae, Eleotridae) pondent en eau douce, les larves dévalent vers la mer où elles vivent une phase planctonique d’une durée très variable (3 à 8 mois selon les espèces). Elles retournent ensuite vers les rivières pour leur croissance et leur reproduction. Ces poissons constituent souvent la plus grande part des peuplements dans les systèmes dulçaquicoles insulaires et montrent les taux d’endémisme les plus élevés. En raison des remontées massives des larves dans les rivières à certaines périodes de l’année, ces espèces sont aussi, dans certains archipels, une ressource alimentaire importante (à l’état larvaire ou adulte) pour les populations locales, mais une ressource très fragile, parfois même au bord de l’extinction, en raison de la complexité de leur cycle biologique et des impacts anthropiques. Compte tenu de leur mode de dispersion (larves marines), l’étude des facteurs qui conduisent à l’endémisme ou à une plus large distribution est primordiale pour la compréhension de la phylogéographie des espèces.
Le massif karstique de Lengguru est un site exceptionnel pour apporter des éléments de réponse. Cette étude s’intégrera dans le programme transversal de l’UMR BOREA et s’appuiera sur son réseau régional (Vanuatu, Nouvelle-Calédonie, Polynésie, etc.).
Les massifs karstiques sont également connus pour abriter des faunes spécifiques de gastéropodes hydrobies, crustacés et insectes aussi bien en surface que dans les milieux souterrains.

L’objectif sera d’inventorier la diversité de ces groupes taxonomiques dans les différentes zones d’étude et d’analyser leur processus de diversification selon une approche similaire à celle développée chez les poissons.
L’hypothèse de barrière à la dispersion des nombreuses disparition-résurgence observées dans les réseaux karstiques et hydrographiques fragmentés sera également testée chez tous les grands groupes d’organismes étudiés.
La datation des évènements géologiques responsables de la fragmentation et de l’isolement des écosystèmes souterrains et de surface permettra éventuellement de proposer une calibration temporelle des taux de divergence moléculaires estimés entre les populations échantillonnées dans chaque type de milieu.
Les données apportées par l’étude de la biodiversité passée et actuelle de Lengguru permettra à terme de comparer les patrons de diversification de différents groupes zoologiques dans un contexte environnemental connu et contraint dans le temps, en fonction de leur valence écologique et leur cycle biologique.
Archéologie : art préhistorique unique
Cette région sud-ouest de la Papouasie Occidentale est peu connue sur un plan archéologique en comparaison avec les découvertes réalisées dans le reste de la Papouasie occidentale ou même en Papouasie centrale (Dani, Baliem, etc.). Les données récentes sur le peuplement préhistorique ancien de la partie Ouest de la Papouasie occidentale ont été mises à jour dans le centre de la Tête de l’Oiseau dans deux sites « chasseurs-cueilleurs » en grotte (Kria Cave et Toe Cave) dont les niveaux les plus anciens remontent à environ 26.000 ans. Les autres dates connues pour la préhistoire de cette partie Est avoisinent les 30.000 ans comme dans les sites de la Baliem ce qui correspond à la chronologie préhistorique connue pour la Papouasie-Nouvelle-Guinée (Kuk, 30.000 ans ; Kosipe, 26.000 ans) où grand nombre de collègues australiens travaillent.
 |
Si aucune chronologie préhistorique n’est pour l’heure connue à Kaimana, Bitsyari et Triton, la richesse en sites à peintures pariétales d’époque préhistorique et historique est inégalée ou presque. En effet, l’existence d’art préhistorique dans la zone sud-ouest de l’Irian Jaya fut signalée pour la première fois en 1678 par Johannes Keyts, un marchand au sud de Fakfak sur des falaises du Golfe de Speelman. A la même époque, un autre rapport mentionne également des falaises rougies de peintures dans la région du Golfe McCluer aujourd’hui appelé Golfe Berau. Plus au sud, la zone qui nous intéresse des trois baies de Kaimana, Bitsyari et Triton est exceptionnellement riche en grottes, abris sous roche et falaises où des dessins, des mains négatives et des symboles ornent les parois calcaires. Ces peintures probablement préhistoriques et historiques restent sans datation.
 |
Les premières recherches et publications sur ce thème dans cette zone datent des années 1940 par W.J. Cator qui décrit avec précision des œuvres du passé en se risquant à des hypothèses sur leurs origines. W.J. Cator va même à conclure que les peintures imposantes de ces trois baies ont été réalisées par les ancêtres des groupes vivant encore dans ces régions. Toutefois, c’est à Roder que l’on doit le plus de détail sur l’art préhistorique. De nombreuses autres publications confirment le potentiel archéologique de ces trois baies. Le principal objectif de cette expédition consistera à établir une chronologie préhistorique pour cette région des trois baies qui s’appuiera sur un programme de datation géochronologique des occupations archéologiques qui seront découvertes à la fouille mais aussi des peinturespariétales. Des enquêtes ethno-archéologiques seront également effectuées pour connaître l’histoire du peuplement récent/ancien de la zone et recueillir des informations à propos des peintures et de l’éventuelle récupération patrimoniale par les villages des alentours.
Zones cibles

Zone 1
Le lac Mbuta se situe à l’extrémité d’un poljé d’une cinquantaine de kilomètres carrés dont une partie occupe le cœur de la seule structure synclinale de la chaine. Son altitude est d’environ 190m. Il est drainé par une rivière qui se perd en arrivant au contact du synclinal de Kembelangan (3°53’10″S à 3°53’16″S ; 134°32’18″E à 134°31’23″E). Après un parcours souterrain d’environ 3 km sous l’ensellement du synclinal, les eaux ressortent vraisemblablement dans ce qui ressemble à une reculée karstique (3°54’45″S ; 134°30’04″E) à une altitude d’un peu plus de 100m. La percée souterraine se situe probablement le long d’une faille normale, à proximité immédiate de dolines de grande ampleur. L’accessibilité est possible par une piste visible sur Google Earth et qui dessert tous les points d’intérêt. Même si elle n’est plus carrossable, les distances sont tout à fait compatibles avec la marche à pied.
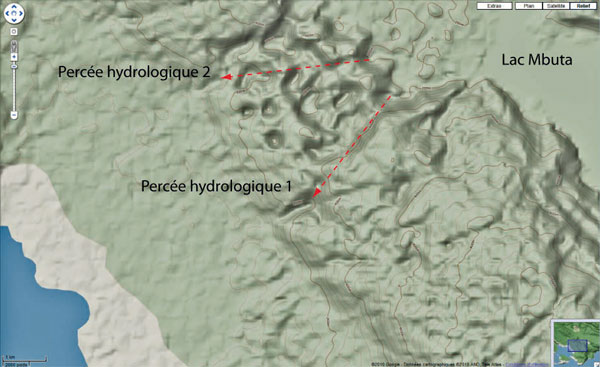
Zone 1 : Percées hydrologiques depuis les
pertes du lac Mbuta en direction de la plaine
côtière à travers l’anticlinal de Kambelanga
trépané par des mégadolines
Zone 2
Les environs du lac Sewiki présentent de multiples intérêts scientifiques. Le lac lui-même est accessible en zodiac via la baie d’Arguni. Il est probablement alimenté par une ou plusieurs résurgences d’eau provenant de l’anticlinal de Berari, mais aussi du lac endoréique situé à moins de 3 km au sud et à une altitude de plus de 300m. Depuis le lac Sewiki, il est également possible d’accéder à peu de distance à un karst d’altitude sur l’anticlinal de Berari. Un affluent de rive gauche de la rivière Berari permet de se rapprocher de grosses dolines sur le flanc Est de l’anticlinal de Seraran.
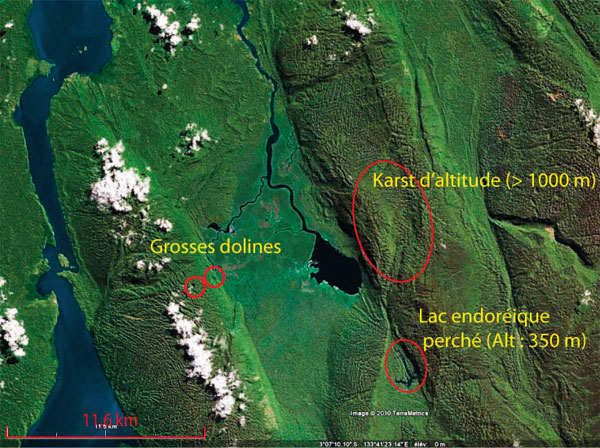
Zone 2 : environs du lac Sewiki
Zone 3
Le poljé de Kuweri se situe à moins de 2 km de la côte au fond de la baie de Bitsyani. La présence proche d’un village laisse envisager une piste d’accès, au moins piétonnière. Une rivière endoréique y circule, il est à moins de 20m d’altitude.

Zone 3 : poljé de Kuweri
Zone 4
La percée hydrologique de la rivière Lengguru qui disparait sur environ 2 km, est d’un intérêt majeur. Il est vraisemblable qu’une piste piétonnière permette de s’en approcher. Il sera nécessaire de s’octroyer l’aide des villageois pour pouvoir atteindre cette zone. En amont de la perte de la Lengguru, des bras morts viennent buter contre ce même relief, ce qui semble indiquer la présence d’anciennes percées hydrologiques, donc des cavités fossiles potentiellement bien plus riches en matériel instructif. Cette perspective est confortée par la présence de porches de cavités importantes observés par Vivien Bailly lors de ses repérages aériens.
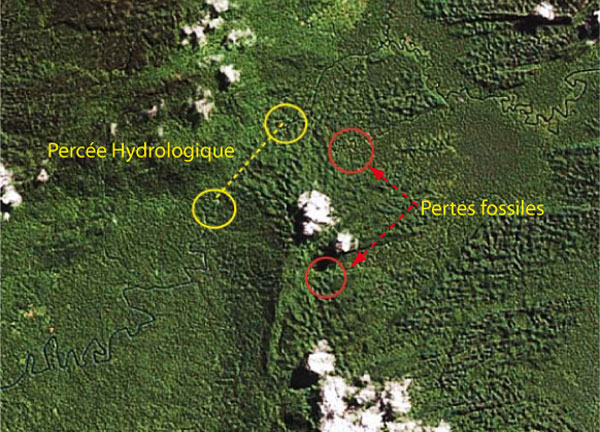
Zone 4 : Percée hydrologique du fleuve Lengguru, et pertes
fossiles supposées à l’aval de bras morts
Zone 5
La région des lacs majeurs situés en arrière de la baie de Triton occupent des poljés étagés entre 100 et 300 m d’altitude. Le lac Kamakawala devrait être assez facilement accessible à pied. Les autres lacs (Lakamora, Aiwasa) et la baie de Triton (recherche de résurgences sous-marines) seront également explorés.

Zone 5 : baie de Triton. Au troisième plan le lac Kamakawallar
(endoréique) et sa frange de battement